
I. Responses of plants to the excess of trace metallic elements
A. Cadmium accumulation and tolerance in plants
Trace metallic elements like cadmium, mercury and lead are not essential and can be toxic for living organisms even at low concentrations. Cadmium is one of the most important environmental pollutants and one of the most toxic heavy metals. Cadmium accumulation in soils originates from atmospheric deposition, application of sewage sludge, phosphate fertilisers, and industrial activities ( from zinc ores mining activities ; cadmium is also a common plastic stabiliser). Cadmium toxicity is thought to be due to its ability to compete with other metals (Ca++ ; Zn++ ; ...) in the proteins as well as high reactivity with thiol groups. (reviewed in Verbruggen et al. 2009a Current opinion in Plant Biology, 12: 364-372)
Accumulation of cadmium by plants is a major route for entry in the food chain. A better understanding of the underlying mechanisms of metal uptake and processing by plants can help to devise remedy to avoid accumulation of toxic heavy metals in crops. On the other hand, the use of plants represents a promising strategy to decontaminate or stabilise polluted soils. This strategy is called phytoremediation. Yet, its development is limited by our current knowledge of the biochemical, physiological and ecological processes involved in metal tolerance and accumulation in plants.
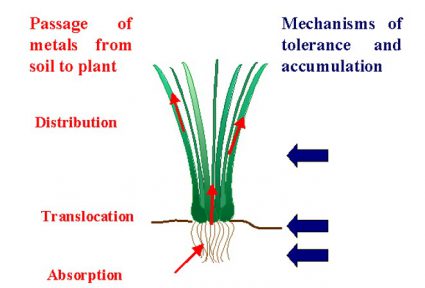
Fig. 1. Overview of our current understanding of adaptations to hyperaccumulate metals. REF Verbruggen et al. 2009b New Phytologist
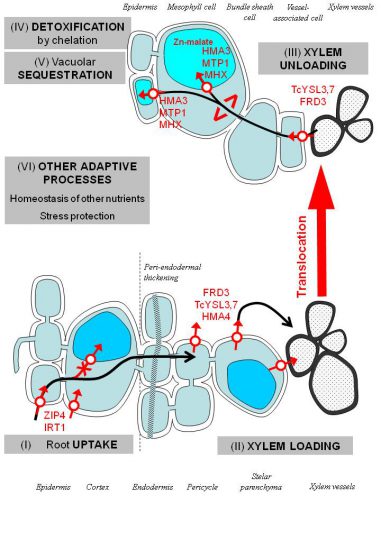

Our model species are Arabidopsis halleri and Noccaea caerulescens (previously named Thlaspi caerulescens), are two Cd hyperaccumulators. Both are close relatives of Arabidopsis thaliana for wich molecular tools have been largely developped. Furthermore this project uses crosses between metallicolous and non metallicolous populations as well as a collection of populations with contrasting tolerance and metal accumulation capacities.
Several strategies are being developed in our laboratory to identify genes involved in Cd tolerance and hyperaccumulation.

Screening of cadmium resistant yeasts
Growth of S. cerevisiae (strain BY474)
Dilution of S. cerevisiae BY474, transformed with empty (pYX212) or recombinant plasmid, spotted onto growth medium containing 20 µM Cd. 3 clones from the Cd screening are presented. Several Noccaea caerulescens genes, TcHMA4, TcMT1, TcMT2, TcMT3, identified in the screen have been further analyzed.
Bernard et al. 2004, Roosens et al. 2004, Roosens et al. 2005, Meyer et al. 2011
Comparison of Noccaea caerulescens or Arabidopsis halleri populations
Populations originating from heavy metal polluted or non polluted (metalliferous) soils, have been studied in collaboration with Prof. A. Smith (University of Oxford). (Roosens et al. 2003)
This study has revealed that the growth of some metallicolous populations (originating from South of France, St Félix-de-Pallière) was enhanced by the addition of Cd concentrations that are toxic for other populations. The molecular mechanism behind this effect is not deciphered yet.

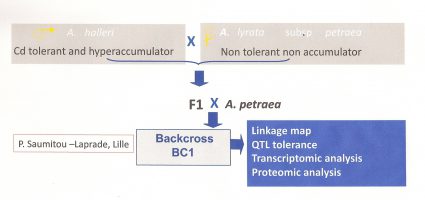
A first genetic study of Cd tolerance was performed by using crosses between A. halleri (Auby population, North of France, Cd and Zn tolerant and hyperaccumulator) and A. lyrata (non tolerant and non hyperaccumulator) (P. Saumitou-Laprade, Lille). A F1 has been back-crossed with A. lyrata. The back-cross progeny has been characterised for Cd tolerance and three quantitative trait loci (QTL) explaining more than 80% of the Cd tolerance character have been identified. (Bert et al. 2003, Willems et al. 2007, Courbot et al. 2007 )
The major QTL for Cd tolerance co-localizes with HMA4 encoding a heavy metal pump, involved in the cellular efflux of Zn++, Cd++ and translocation of those metals from the root to the shoot (Courbot et al. 2007).
The second major QTL for Cd tolerance co-localizes with CAX1 encoding a Ca++/H+vacuolar antiporter (Baliardini et al. 2015). CAX1 seems to play an important role in Cd tolerance of plants by limiting oxidative stress. CAX1 activity is thought to disrupt the positive feedback between increasy in cytosolic Ca++ and ROS (Baliardini et al. 2015, Baliardini et al. 2015).
Using the same strategy with a Arabidopsis halleri non metallicolous population the QTL analysis revealed only HMA4 as sole major determinant for the Cd and Zn tolerance traits. This result provides evidence that preadaptation process in Arabidopsis halleri prior colonization of Zn and Cd contaminated sites included HMA4 duplication (Meyer et al. 2016)
A F2 population was also characterized. QTL analysis for Cd accumulation allowed the identification of one locus explaining 20% of the Cd accumulation character and many loci with small additional effects (Willems et al. 2010).
Overexpression of selected genes in Arabidopsis thaliana
Selected genes are overexpressed in Arabidopsis thaliana, which is the easiest transformable plant since transformation can be done by flower dipping. As A. thaliana is very sensitive to cadmium, effect of gene overexpression on Cd tolerance can be easily measured. If enhanced cadmium accumulation and/or tolerance is observed, application will be foreseen in cultivated species.


Overexpression of selected Noccaea caerulescens cDNAs in Nicotiana plumbaginifolia
Selected genes are overexpressed in Nicotiana plumbaginifolia, which is a high biomass easy transformable species. Better growth on contamined soils of Brussels could be observed for some lines (Alban Heudiard).

B. Copper accumulation and tolerance in plants
We have started a research on copper tolerance within the PIC project “Remedlu” in Katanga (RD Congo) coordinated by Pierre Meerts (ULB). The so-called copper arc in Katanga and Zambia (Central Africa) is one of the most mineralized regions in the world. In particular copper in soil can be as high as 50.000 ppm while normal soils contain around 10 ppm – 25 ppm. The flora is there unique, characterized by a high degree of endemism.
We have studied Haumaniastrum katangense, called the copper flower, which shows extreme tolerance to copper and high accumulation capacity (PhD thesis of François Chipeng, Chipeng et al. 2009). We have demonstrated a higher requirement for copper for normal growth. Surplus copper was also required for cultivating H. katangense in sterile conditions, suggesting that Cu excess may be necessary for needs other than pathogen defence. Further studies have included the presence of cobalt excess in the soils of the copper arc. In particular, the endemic species and obligate metallophyte Haumaniastrum robertii was studied together with its facultative metallophyte congener H. katangense. H. robertii growth was conditional on addition of both copper and cobalt. Furthermore the beneficial effect of cobalt on the plant growth was conditional on simultaneous copper supplement. Co and Cu accumulate in different tissues of Haumaniastrum and they show different mobility in plants (PhD thesis of Francine Ilunga Kabeya, Kabeya et al. submitted)
The endophytic community of H. katangense and Crepidorhopalon tenuis, an other cuprophyte of the copper arc, was studied in collaboration with the Institut de Recherches Microbiologiques JMW/Laboratoire de Microbiologie and Max Mergeay (CEN/SCK), as a first step to evaluate their potential contribution to plant adaptation to copper excess (PhD thesis of Alfred Cubaka ; Cubaka et al. 2010). Bacteria with outstanding resistance to copper have been identified in the endophytes of both cuprophytes and in their rhizosphere (manuscript in preparation).
II. Response of plants to nutrient deficiency
A. Contribution to the study of magnesium homeostasis
- Plant nutrition
Understanding how plants regulate absorption, transport and allocation of nutrients can have major applications for the environment and human health. By improving those processes, it is possible to develop crops that grow more efficiently on nutrient-poor soils (reduced fertilizers input) and crop products with added nutritional value (biofortification). Two research themes are focused on plant nutrition : magnesium homeostasis and the impact of nitrogen availability on the root system in the model species Arabidopsis thaliana. On the one hand, the aim is to understand how plants acquire and regulate Mg content internally. Physiological and transcriptomic approaches have identified transport of sugars and the circadian clock as early targets of Mg deficiency. On the other hand, the mechanisms that govern the plasticity of the root system in response to nitrate are studied (low concentrations stimulate the development of lateral roots, while consistent high levels have an inhibitory effect). We use genetic approaches to exploit natural variability in populations of Arabidopsis to identify genes that regulate root architecture. For those two lines of research, knowledge transfer is considered to agronomic species of the Brassica genus (cabbage, oilseed rape ...).
- Magnesium
Magnesium is a priority metal because in addition to being an essential element in plant cell biology, it is the 4th common cation in the human body and half of its dietary intake is of plant origin. Hypomagnesaemia is recognized as a global clinical problem.
- Physiological characterization of Mg deficiency in plants
Magnesium deficiency in plants is a widespread problem, affecting productivity and quality in agriculture, horticulture and forestry. In arable land, the incidence of Mg deficiency symptoms is increasing, due to intensive harvesting, amplified rotations per site and unbalanced application of fertilizers lacking in secondary elements (e.g. calcium, magnesium).
The targets of Mg deficiency on the photosynthetic apparatus were studied in sugar beet: the two photosystems showed sharply contrasting responses during the acclimation process to low Mg supply (Hermans et al. 2005a). Photosystem II was down-regulated through a loss of antenna, whereas photosystem I was most probably down-regulated through a more important loss of reaction centres. As sugars accumulated in leaves before any loss of photosynthetic activity, we also examined the impact of low Mg status on sugar partitioning. We showed that sucrose export was an early target of Mg deficiency in Arabidopsis (Hermans et al. 2005b) and sugar beet (Hermans et al. 2005a). Interesting findings were that in response to low Mg level, a sucrose/H+ symporter gene was induced, however without further enhancement of the protein level and the sucrose loading into the phloem. Finally, we also showed that deficiencies of other major elements (N, P, K) can be used to manipulate sugar export and to alter biomass allocation (Hermans et al. 2006).
- Magnesium homeostasis in plants and interaction with the circadian clock
Two approaches are used to expand knowledge on the mechanisms underlying Mg homeostasis in Arabidopsis, a domain that was relatively unexplored to that point. The research focus is on (i) the exploitation of ionomic variation and (ii) the identification of transcriptome changes related to Mg limitation.
(i) The first approach uses the variation of Mg concentration in Arabidopsis mutants and natural accessions as a source of diversity to find new genes and alleles controlling root and shoot Mg homeostasis.
(ii) Because the knowledge about the impact of Mg deficiency on physiological processes was scarce, we proceeded to transcriptome analyses to provide non-biased hints about targets of Mg starvation. We published a thorough description of the transcriptomic responses (within hours or days before the outbreak of visual symptoms) of Mg deprivation (Hermans et al. 2010a & b). Unlike other mineral deficiencies, putative Mg transporters genes were not induced (post-transcriptional induction cannot be excluded). Interestingly, the rhythmic expression of circadian clock component genes was altered in roots, while abscisic acid signalling was triggered in leaves. The transcriptomic analysis confirmed the visual observation that Mg starvation affected the shoot more than the root in Arabidopsis. Further study of the circadian rhythm confirmed interaction between the circadian clock and Mg status. This aspect is currently further investigated.
Effects of Mg deficiency on photosystems
Visual symptoms of Mg deficiency in sugar beet. Mg deficiency appears first on the most recently developped leaves as chlorosis between the veins, which remind green. Brown spottings and necroses appear when deficiency is severe or under high light intensity.

The behaviour of PSII and PSI was assessed using direct and modulated fluorescence measurement, near-infrared absorbance changes and 77K fluorescence emission spectroscopy. An early effect on the maximum PSI oxidation rate could be identified before any decrease in the rate of PSII maximum quantum yield of primary photochemistry, PSII electron transport or any chlorophyll degradation. Also, a decrease in the amplitude fluorescence emission peak at 735 nm has been observed, suggesting an early effect of Mg deficiency on PSI. Concomitantly, sucrose accumulates in source leaves upon Mg deficiency as an early response. High sucrose levels are known to down-regulate the chloroplast electron chain transport. The delay of photosynthesis regulation in Mg-deficient plants may provoke an unbalance between light and dark reactions, so inducing oxidative stress and further leading to necrogenesis on leaves.
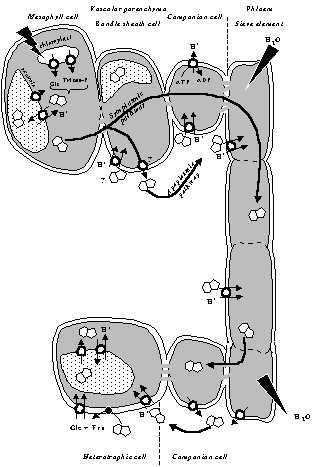
Fig. from Hermans et al. 2006
Effects of Mg deficiency on sucrose partitioning
Sucrose pathway from production to consumption and storage organs. In sugar beet, sucrose loading from the apoplasm to companion cell is operated trough sucrose/H+ symporter.
Impact of magnesium deficiency on plant metabolism is ill known. The adaptation mechanisms of the primary photochemical apparatus to the lack of magnesium is not the first target of magnesium deficiency. Although Mg is found at the centre of the tetrapyrol ring, the lowering in chlorophyll, as a result of the lack of this element could only be established after a decrease in assimilation rate had already occured. A major effect of magnesium deficiency seems to be inhibition of sucrose translocation. More recent cellular targets have been recently identified by a global transcriptonics studied (Hermans et al. a,b).
Comments are closed, but trackbacks and pingbacks are open.